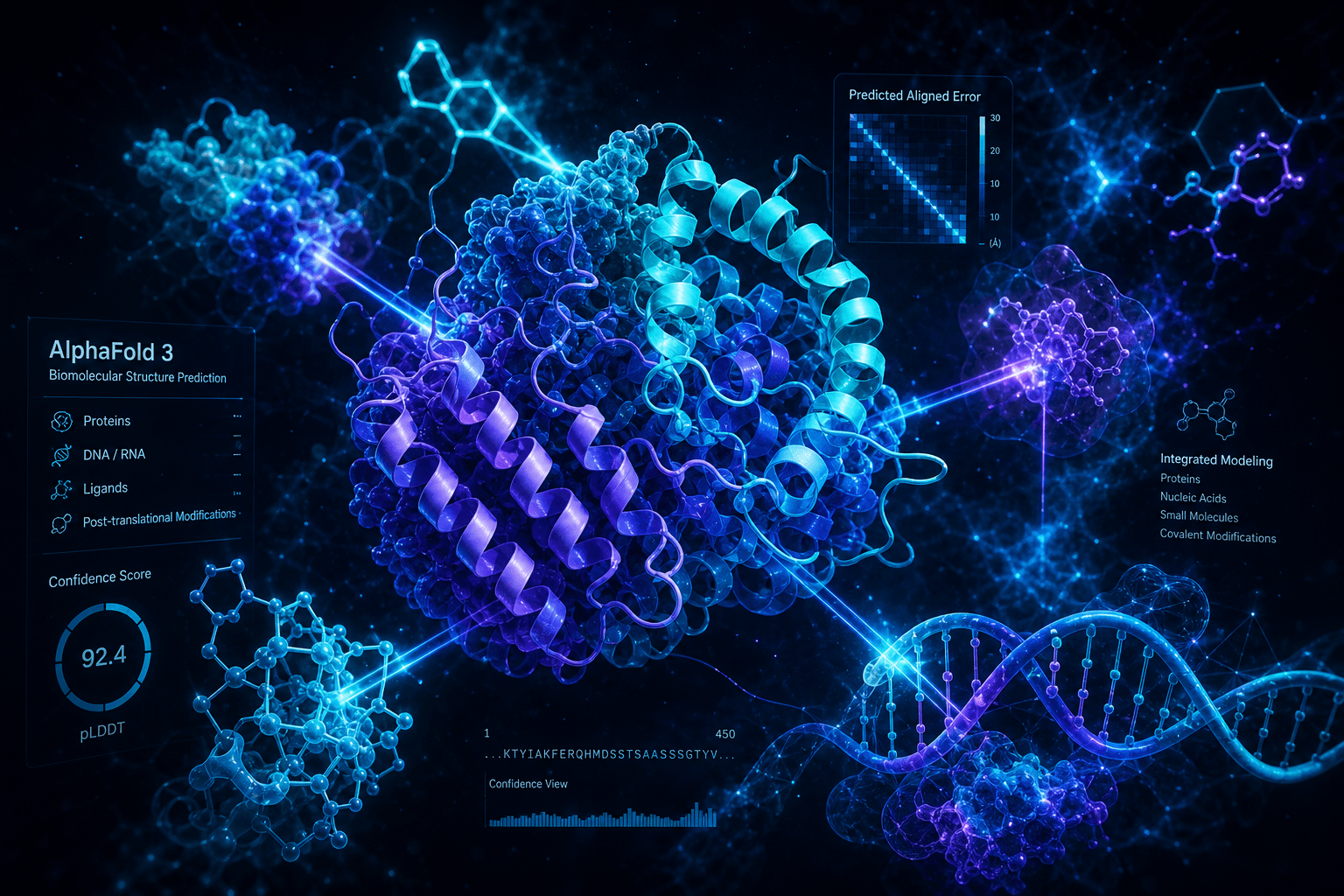
AlphaFold 3 in Nature: DeepMind's Free Server and the Biomolecular Revolution

Table of Contents
AlphaFold 3 in Nature: DeepMind's Free Server and the Biomolecular Revolution #
AlphaFold 3 publishes in Nature today with a free server now available for researchers worldwide. This is not an incremental update—it is a fundamental expansion of what AI can predict about life at the molecular level.
For the first time, a single unified model accurately predicts the structure and interactions of nearly every molecule type found in the Protein Data Bank: proteins, DNA, RNA, small molecule ligands, ions, and post-translational modifications. The implications ripple through drug discovery, structural biology, and our fundamental understanding of cellular machinery.
DeepMind and Isomorphic Labs have simultaneously launched the AlphaFold Server at alphafoldserver.com—free for non-commercial research use—putting this capability in the hands of scientists who previously needed months of crystallography work or expensive specialized software to approximate what AlphaFold 3 generates in minutes.
This is the realization of a broader vision: moving from predicting individual protein structures to modeling complete biomolecular systems with all their interacting parts. The technical architecture underpinning this shift replaces AlphaFold 2's Evoformer and structure module with a streamlined Pairformer and a diffusion-based structure generator—changes that enable the model to handle arbitrary chemical compositions without specialized handling for each molecular type.
Table of Contents #
- What AlphaFold 3 Does Differently: Beyond Proteins to All Biomolecules
- From Protein-Only to Universal Biomolecular Prediction
- The Biomolecular Interaction Problem AlphaFold 3 Solves
- The Architecture Shift: How AlphaFold 3 Works Under the Hood
- Pairformer and Diffusion Heads Replace Evoformer
- Confidence Metrics That Actually Matter
- The Free AlphaFold Server: What Just Went Live
- Server Capabilities and Limitations
- How to Access It Today
- Implications for Drug Discovery and Pharmaceutical Research
- Small Molecule and Antibody Prediction
- Competitive Pressure on Traditional Methods
- Extending the AlphaFold Legacy: DeepMind's Broader Vision
- The Isomorphic Labs Connection
- From Open Database to Active Research Platform
- Performance Benchmarks: What the Numbers Actually Show
- CASP15 and Recent Evaluations
- Where AlphaFold 3 Still Struggles
- Technical Limitations and Honest Caveats
- Dynamic Conformations and Predictive Uncertainty
- The Training Data Question
- What This Means for AI in Life Sciences
- Democratization of Structural Biology
- The Next Research Frontier
What AlphaFold 3 Does Differently: Beyond Proteins to All Biomolecules #
AlphaFold 3 expands the scope of AI structure prediction from single proteins to complete biomolecular systems. While AlphaFold 2 transformed protein structure prediction, it treated proteins largely as isolated entities. AlphaFold 3 models how proteins interact with DNA, RNA, small molecules, and each other in a single unified framework.
The practical difference is substantial. A researcher studying a transcription factor no longer needs separate tools for predicting the protein structure, docking it to DNA, and modeling ligand binding. AlphaFold 3 accepts a combined input—protein sequences, nucleic acid sequences, and small molecule SMILES strings—and generates a unified 3D structure showing how all components fit together.
The Nature paper demonstrates this capability across multiple biologically critical systems:
| Complex Type | Example | Biological Significance |
|---|---|---|
| Protein-DNA | CRP/FNR family transcriptional regulator with cGMP | Gene regulation mechanisms |
| Protein-RNA | 40S ribosomal subunit with initiation factors | Translation initiation |
| Protein-ligand | LGK974 clinical inhibitor bound to PORCN | Drug target validation |
| Antibody-antigen | Mesothelin-targeting therapeutic antibody | Biologic drug development |
| Glycoprotein | EXTL3 homodimer with glycosylation | Protein modification effects |
This unified approach matters because biology does not operate in isolated molecular types. Enzymes bind substrates (small molecules). Transcription factors recognize DNA sequences. Antibodies neutralize viral proteins. AlphaFold 3 captures these interactions natively rather than requiring researchers to stitch together predictions from disparate specialized tools.
From Protein-Only to Universal Biomolecular Prediction #
The input interface has been generalized to accept virtually any biomolecular entity. AlphaFold 3 processes polymer sequences (proteins, DNA, RNA), residue modifications, and ligand chemical specifications through a unified tokenization scheme that treats all molecular types as first-class citizens.
The tokenization is dynamic—amino acids can be represented at different scales depending on context, from backbone-only coordinates to full side-chain representations. This flexibility allows the model to handle the combinatorial explosion of possible molecular compositions without requiring separate architectural paths for each type.
Key molecular types now supported include:
- Standard amino acids and modified protein residues (phosphorylation, glycosylation, methylation)
- DNA and RNA in single-stranded and double-stranded conformations
- Small molecule ligands specified by SMILES strings
- Metal ions and other atomic entities
- Covalent modifications including bonded ligands and glycans
The training data reflects this breadth. The model was trained on structures from the Protein Data Bank released before September 30, 2021, with careful filtering to ensure diverse coverage across molecular types. Unlike previous specialized predictors that trained on narrow subsets (protein-ligand dockers trained only on bound complexes, RNA predictors trained only on RNA structures), AlphaFold 3 learns from the full diversity of deposited structures.
The Biomolecular Interaction Problem AlphaFold 3 Solves #
Predicting how molecules interact has historically required specialized tools with limited generalization. Protein-ligand docking tools like AutoDock Vina require pre-defined protein pockets and struggle with induced fit. Protein-DNA predictors like RoseTTAFold2NA handle only specific interaction types. Antibody-antigen prediction remained a distinct subfield with its own methods.
AlphaFold 3 breaks down these silos. The Nature paper reports accuracy improvements that directly challenge the specialized state of the art:
| Interaction Type | Improvement vs. Specialized Tools | Significance |
|---|---|---|
| Protein-ligand | Far greater accuracy vs. docking tools (P < 2.27 × 10−13) | Replaces physics-based docking for hit identification |
| Protein-nucleic acid | Higher accuracy vs. nucleic-acid-specific predictors | Enables transcription factor-DNA modeling |
| Antibody-antigen | Substantially higher accuracy vs. AlphaFold-Multimer v2.3 (P = 6.54 × 10−5) | Accelerates therapeutic antibody development |
| Covalent modifications | Successful prediction of 76.6% of bonded ligands | Captures post-translational regulation |
The protein-ligand results are particularly notable for drug discovery. On the PoseBusters benchmark—428 recent protein-ligand structures not in the training data—AlphaFold 3 achieves accurate pocket-aligned predictions on a significantly higher percentage of targets than traditional docking tools, even though those tools receive privileged information about the binding pocket that AlphaFold 3 does not use.
This matters because drug discovery is fundamentally about molecular interactions. A small molecule drug works by binding to a protein target. A CRISPR guide RNA works by recognizing a specific DNA sequence. An antibody therapy works by locking onto a viral surface protein. AlphaFold 3 models these interactions directly, potentially compressing months of experimental validation into minutes of computation.
The Architecture Shift: How AlphaFold 3 Works Under the Hood #
The architecture represents a substantial evolution from AlphaFold 2, with three major changes: the Pairformer module replaces the Evoformer for processing, a diffusion-based structure module replaces the IPA-driven structure generation, and confidence estimation now handles generative outputs. These changes are necessary to accommodate arbitrary chemical compositions without specialized handling for each molecular type.
The overall flow remains conceptually similar: an input processing stage, a large trunk network that evolves pairwise representations, and a structure module that generates explicit atomic coordinates. But each stage has been re-engineered for generality and data efficiency.
AlphaFold 3 Architecture Flow:
Input → Tokenization → MSA Embedding → Pairformer (48 blocks) →
Diffusion Module → All-atom coordinates + Confidence metrics
Key differences from AF2:
- Pairformer replaces Evoformer (simpler, more efficient)
- Diffusion module replaces structure module (generates arbitrary molecules)
- Confidence head adapted for generative outputsThe MSA (Multiple Sequence Alignment) processing has been substantially de-emphasized. Where AlphaFold 2 used a heavy Evoformer with both sequence and pair attention, AlphaFold 3 reduces MSA processing to a minimal embedding step followed by pair-weighted averaging. The MSA representation does not persist through the trunk—only the pair representation carries information forward. This simplification proves that massive MSA processing, while important for AlphaFold 2's protein-only predictions, is not necessary for general biomolecular modeling.
Pairformer and Diffusion Heads Replace Evoformer #
The Pairformer is the computational heart of AlphaFold 3—a simplified but powerful replacement for the Evoformer. Where the Evoformer maintained both an MSA representation and a pair representation throughout 48 processing blocks, the Pairformer operates only on the pair representation and a single representation, with MSA information distilled early through pair-weighted averaging.
This architectural choice reflects a key finding: for general biomolecular prediction, the evolutionary information captured in MSAs is less critical than the geometric constraints encoded in the pair representation. The Pairformer processes these constraints through 48 blocks of attention and transition operations, outputting refined pair and single representations that feed into the structure module.
The structure module is where the most dramatic change occurs. AlphaFold 2 used Invariant Point Attention (IPA) operating on backbone frames and side-chain torsion angles—a representation inherently tied to protein chemistry. This worked brilliantly for proteins but could not generalize to arbitrary molecules.
AlphaFold 3 replaces this with a diffusion module that operates directly on raw atom coordinates. The approach is conceptually straightforward:
- Forward process: During training, noise is added to true atomic coordinates to create "noised" structures
- Learning objective: The network learns to predict the true coordinates from noised versions
- Inference process: Random noise is sampled and iteratively denoised to generate final structures
The diffusion process operates at multiple scales simultaneously. At low noise levels, the network focuses on local stereochemistry—bond lengths, angles, and local geometry. At high noise levels, it learns global structure—the overall fold and domain arrangement. This multiscale nature eliminates the need for the complex torsion-based parameterizations and stereochemical violation losses that AlphaFold 2 required.
Critically, this is a generative model. Unlike AlphaFold 2, which produced a single deterministic output, AlphaFold 3 can generate multiple plausible conformations by running the diffusion process with different random seeds. For antibody-antigen prediction—where the binding interface can adopt multiple conformations—running 1,000 seeds and ranking by confidence produces significantly better results than single-seed prediction.
Confidence Metrics That Actually Matter #
AlphaFold 3 provides calibrated confidence metrics that indicate which predictions to trust and which to treat skeptically. These metrics—pLDDT, pTM, and ipTM—are familiar from AlphaFold 2 but required significant adaptation for the diffusion-based architecture.
The challenge is that diffusion training only supervises a single denoising step, not the full structure generation process. To enable confidence prediction, the researchers developed a diffusion rollout procedure during training: the model generates full structures using larger step sizes, then the confidence head learns to predict the accuracy of these rollouts.
The resulting confidence measures are:
| Metric | Measures | Range | When High Values Indicate Trust |
|---|---|---|---|
| pLDDT | Per-residue local distance difference test | 0-100 | High (90+) for structured regions, low (<50) for disordered tails |
| pTM | Predicted TM-score for entire complex | 0-1 | Values > 0.8 suggest overall correct fold |
| ipTM | Interface-predicted TM-score | 0-1 | Critical for interactions; > 0.8 indicates reliable interface prediction |
The calibration data in the Nature paper shows these metrics track actual accuracy remarkably well. When ipTM scores are high, protein-protein interfaces are genuinely accurate (high DockQ). When pLDDT scores drop below 50, the corresponding regions are typically unstructured in reality.
This calibration enables practical decision-making. A researcher can run a prediction, check the confidence metrics, and decide whether the result is reliable enough for their application—or whether they need to run additional seeds, adjust inputs, or pursue experimental validation.
For antibody-antigen prediction specifically, the paper documents that interface quality continues improving even with 1,000 model seeds, significantly beyond the standard 5 seeds used for most predictions. This suggests that high-stakes applications should invest in extensive sampling and confidence-based ranking.
The Free AlphaFold Server: What Just Went Live #
The AlphaFold Server at alphafoldserver.com is live today, offering free access to AlphaFold 3's core capabilities. This is a strategic move by DeepMind and Isomorphic Labs—democratizing access while establishing the infrastructure for a broader biomolecular research platform.
The server interface is deliberately simple. Researchers paste sequences or enter small molecule names, and the system returns structure predictions within minutes. No installation, no GPU clusters, no wrestling with dependencies—just a web form and results.
Since launch, researchers have already generated over 1 million structures through the platform. The usage pattern suggests immediate adoption across the research community.
However, the server comes with important restrictions that define its current scope:
| Aspect | Server Offering | Limitations |
|---|---|---|
| Cost | Free for non-commercial research | Commercial use prohibited |
| Input complexity | Limited number of tokens | Large complexes may not fit |
| Ligand selection | Curated set of biologically common molecules | Custom small molecules restricted |
| Output usage | Visual analysis, publication | Cannot use for docking/screening tool training |
| API access | Web interface only | No programmatic batch processing |
These restrictions are designed to enable broad scientific access while protecting Isomorphic Labs' commercial drug discovery interests and preventing model distillation by competitors.
Server Capabilities and Limitations #
The AlphaFold Server supports the most common biomolecular prediction scenarios while excluding high-complexity commercial applications. Understanding these boundaries helps researchers set appropriate expectations.
Capabilities include:
- Protein-only predictions (standard AlphaFold 2-equivalent functionality)
- Protein-protein complexes including antibody-antigen interactions
- Protein-DNA and protein-RNA complexes for transcription factor and ribosome modeling
- Limited protein-ligand predictions using a curated ligand library
- Glycoprotein predictions with common glycosylation patterns
- Covalent modifications including phosphorylated residues and bonded ligands
Key limitations researchers should be aware of:
Token limits: The server enforces maximum sequence lengths and total complex sizes. Large ribosomal assemblies or multi-protein mega-complexes may not fit.
Ligand restrictions: While the Nature paper demonstrates prediction of arbitrary small molecules via SMILES strings, the server limits ligands to a predefined set of biologically relevant molecules. Custom drug-like compounds are not currently supported in the free interface.
No batch processing: The web interface is designed for interactive exploration, not high-throughput screening. Researchers needing hundreds or thousands of predictions must wait for academic code release or build their own inference infrastructure.
Usage restrictions: The terms of service explicitly prohibit using outputs to train competing machine learning models or develop docking/screening tools. This protects DeepMind and Isomorphic Labs' investment while enabling academic research.
How to Access It Today #
Accessing AlphaFold 3 through the server requires only a Google account and a web browser. Navigate to alphafoldserver.com, sign in, and start a new prediction.
The input interface accepts:
Example Input Configuration:
Protein 1: >sp|P01308|INS_HUMAN
MALWMRLLPLLALLALWGPDPAAAFVNQHLCGSHLVEALYLVCGERGFFYTPKTRREAEDL
QVGQVELGGGPGAGSLQPLALEGSLQKRGIVEQCCTSICSLYQLENYCN
Protein 2: [optional second chain]
DNA: CTGACTGATCGATCGTAGCTAGCTAGCTAGCTAGCTAGCTAGCT
Ligand: [select from curated list]Typical runtimes range from 2-10 minutes depending on complex size. The system automatically handles MSA generation, template search, and structure prediction. Results include:
- 3D structure visualization in an interactive molecular viewer
- Confidence scores (pLDDT, pTM, ipTM) for each chain and interface
- Predicted Aligned Error (PAE) matrix showing per-residue confidence relationships
- Downloadable structure files in mmCIF format for downstream analysis
For researchers needing programmatic access or custom ligand support, DeepMind has indicated that inference code will be released for academic use—though without model weights for the full system. This partial release enables method development and validation while maintaining competitive control over the most capable version.
Implications for Drug Discovery and Pharmaceutical Research #
AlphaFold 3 changes the economics of structure-based drug design. Historically, obtaining a high-quality structure of a protein target required months of crystallization work or expensive cryo-EM analysis—often costing tens of thousands of dollars and delaying programs. AlphaFold 3 generates comparable structural information in minutes at negligible marginal cost.
The implications cascade through the drug discovery pipeline:
| Stage | Traditional Approach | AlphaFold 3-Enabled Approach |
|---|---|---|
| Target validation | Literature mining, expression screening | Immediate structure prediction for any sequence |
| Hit identification | High-throughput screening (HTS) | Structure-based virtual screening with predicted complexes |
| Lead optimization | Iterative synthesis and assay | In silico prediction of ligand binding before synthesis |
| Selectivity assessment | Cross-screening assays | Predictive modeling of off-target binding |
| Biologics design | Phage display, hybridoma generation | Computational antibody optimization against predicted epitopes |
The financial stakes are substantial. Isomorphic Labs—the DeepMind spinout applying AlphaFold technology to commercial drug discovery—announced two major partnerships in January 2024: a $45 million upfront deal with Eli Lilly (worth up to $1.7 billion with milestones) and a $37.5 million upfront deal with Novartis (worth up to $1.2 billion). These agreements include tiered royalties on net sales and represent a vote of confidence from major pharmaceutical companies that AlphaFold-derived technology can meaningfully accelerate therapeutic development.
The technology is not just theoretical. The Nature paper demonstrates accurate prediction of clinically relevant systems: the structure of LGK974, a clinical-stage cancer inhibitor, bound to its PORCN target protein; the interaction of therapeutic antibodies with viral spike proteins; and the recognition of DNA by transcription factors that regulate gene expression.
Small Molecule and Antibody Prediction #
The protein-ligand and antibody-antigen capabilities are the most commercially relevant features. These represent the core modalities of modern drug discovery—small molecule oral drugs and injectable biologics—and the areas where AlphaFold 3 demonstrates the strongest performance improvements.
For small molecules, AlphaFold 3 achieves what the Nature paper calls "far greater accuracy for protein-ligand interactions compared with state-of-the-art docking tools." The PoseBusters benchmark comparison is striking: AlphaFold 3 succeeds on significantly more targets than classical tools like Vina and Gold, even though those tools receive privileged pocket information that AlphaFold 3 does not use.
The mechanism is fundamentally different. Traditional docking requires:
- A known or predicted protein structure
- Definition of the binding pocket location
- Sampling of ligand conformations within that pocket
- Scoring based on physics-based or learned energy functions
AlphaFold 3 instead learns the joint distribution of protein and ligand coordinates directly from data. Given a protein sequence and ligand SMILES, it generates the bound complex without explicit pocket specification or conformational sampling. The diffusion process inherently captures both protein flexibility (induced fit) and ligand conformational preferences.
For antibodies, the improvements are equally significant. AlphaFold 3 shows "substantially higher antibody-antigen prediction accuracy compared with AlphaFold-Multimer v.2.3" (P = 6.54 × 10−5). This matters because therapeutic antibody development traditionally requires extensive experimental screening to identify binders with the right specificity, affinity, and developability properties. Accurate computational prediction could prioritize candidates before wet-lab work begins, compressing timelines and reducing costs.
However, the paper notes that antibody prediction quality continues improving with up to 1,000 model seeds—far more than the standard 5 seeds. This suggests that for high-value therapeutic programs, extensive computational sampling remains worthwhile.
Competitive Pressure on Traditional Methods #
AlphaFold 3 applies competitive pressure across the structural biology and computational chemistry landscape. As prediction accuracy improves, the value proposition of experimental determination changes—crystallography and cryo-EM remain essential for validation and novel fold discovery, but their role in routine structure determination diminishes.
The business implications are immediate:
- Structural biology CROs must pivot from routine structure determination to challenging cases (novel folds, dynamic systems, membrane proteins) and validation services
- Computational chemistry software vendors face disruption of their docking and scoring business models; physics-based methods must demonstrate value beyond what learned models provide
- Cryo-EM service providers see demand shift from single-particle analysis of soluble proteins to more complex applications—tomography, in situ structural biology, and time-resolved studies
The Nature paper is careful to note limitations that preserve experimental relevance. AlphaFold 3 does not predict dynamic conformational ensembles—it generates static structures similar to those in the PDB. This means experiments that capture motion, conformational switching, and allosteric regulation remain essential.
Similarly, the model struggles with systems that adopt fundamentally different conformations depending on binding state. The paper documents that E3 ubiquitin ligases, which adopt open conformations when unbound and closed when ligand-bound, are predicted exclusively in the closed state by AlphaFold 3 regardless of input specification. Capturing these state-dependent conformations requires experimental approaches or specialized sampling methods beyond standard AlphaFold 3 inference.
The competitive dynamic is not elimination but complementarity. AlphaFold 3 handles the routine prediction tasks that previously consumed experimental resources, freeing those resources for the frontier problems where experiments remain essential.
Extending the AlphaFold Legacy: DeepMind's Broader Vision #
AlphaFold 3 represents the culmination of a multi-year research program that has transformed structural biology. The trajectory from AlphaFold 2 (2020) through the AlphaFold Database release (2021) to today's unified biomolecular model demonstrates DeepMind's commitment to applying artificial intelligence to fundamental scientific challenges.
The progression tells a clear story:
| Milestone | Year | Significance |
|---|---|---|
| AlphaFold 2 CASP victory | 2020 | Demonstrated that learned models could solve protein folding |
| AlphaFold 2 publication | 2021 | Established the architecture that influenced the entire field |
| AlphaFold Database release | 2021 | Open-sourced 200M+ structures, democratizing access |
| AlphaFold-Multimer | 2022 | Extended to protein-protein interactions |
| AlphaFold 3 announcement | 2024 | Unified biomolecular prediction in a single model |
| AlphaFold Server launch | 2024 | Active research platform, not just static database |
The shift from static database to active prediction platform is strategically significant. The AlphaFold Database provided pre-computed structures for known protein sequences—a valuable but limited resource. The AlphaFold Server enables predictions for novel sequences, custom complexes, and researcher-specific scenarios. It transforms AlphaFold from a reference resource into an active tool in the research workflow.
The competitive landscape has also evolved. Meta's ESMFold demonstrated that protein language models could achieve competitive structure prediction without MSA processing—though at lower accuracy than AlphaFold 2. RoseTTAFold and its derivatives from David Baker's group provided open alternatives. AlphaFold 3 reasserts DeepMind's leadership by extending beyond proteins to the full biomolecular context where language models and specialized predictors struggle to compete.
The Isomorphic Labs Connection #
Isomorphic Labs is the commercial bridge between DeepMind's research and pharmaceutical application. Spun out from DeepMind with Demis Hassabis serving as CEO of both entities, Isomorphic Labs applies AlphaFold technology and complementary AI models to drug discovery partnerships with major pharmaceutical companies.
The business model is clear: DeepMind publishes foundational research and maintains open resources (the AlphaFold Server, the database, selected code releases), while Isomorphic Labs commercializes the technology for drug discovery applications. This structure enables scientific impact while capturing commercial value.
The January 2024 partnership announcements validate this approach:
| Partner | Upfront Payment | Milestone Potential | Focus Area |
|---|---|---|---|
| Eli Lilly | $45 million | Up to $1.7 billion | Small molecule therapeutics, multiple targets |
| Novartis | $37.5 million | Up to $1.2 billion | Small molecules, three undisclosed targets |
Both deals include tiered royalties on net sales, suggesting Isomorphic Labs expects its AI-driven discovery process to yield approved therapeutics. The total potential value—nearly $3 billion across just two partnerships—indicates the pharmaceutical industry's confidence that AlphaFold-derived technology can materially improve discovery success rates.
Isomorphic Labs' technical approach extends beyond AlphaFold 3 alone. The company develops proprietary AI models for chemistry, biology prediction, and disease understanding that integrate with the structural predictions. This broader AI platform, combined with AlphaFold 3's biomolecular capabilities, forms the basis of their "rational drug design" offering.
The Nature paper reflects this dual-purpose development. The research was conducted jointly by DeepMind and Isomorphic Labs scientists, with the free AlphaFold Server maintaining the open science commitment while the full capabilities—including arbitrary ligand support—feed into Isomorphic's commercial platform.
From Open Database to Active Research Platform #
The AlphaFold project has evolved through three distinct phases of access and utility. Understanding this progression clarifies what AlphaFold 3 represents for researchers.
Phase 1: The Original AlphaFold 2 System (2021)
Released as open-source code that researchers could install and run locally. Required significant computational resources—GPUs with substantial VRAM, extensive storage for genetic databases, and technical expertise to configure the pipeline. High barrier to entry but full capability for those who could deploy it.
Phase 2: The AlphaFold Database (2021-ongoing)
Pre-computed structures for approximately 200 million protein sequences across the tree of life. Massive democratization—any researcher could download structures for their protein of interest without computational investment. But limited to pre-computed sequences; novel sequences, custom complexes, and specific ligands were not covered.
Phase 3: The AlphaFold Server (2024)
Active prediction platform for arbitrary inputs. Researchers specify their sequences and molecules of interest; the system generates predictions on demand. Brings together the accessibility of the database with the flexibility of the original code—without requiring local infrastructure.
This progression reflects a broader pattern in AI infrastructure: research systems democratize through managed services. Just as OpenAI's GPT models moved from API-only to ChatGPT's accessible interface, and as image generation moved from Colab notebooks to Midjourney and DALL-E, AlphaFold has moved from code release to database to interactive platform.
The strategic implication is that DeepMind sees the AlphaFold Server not as a temporary offering but as the primary interface for non-commercial researchers going forward. The managed service model enables ongoing model improvements, usage monitoring, and quality control that distributed open-source deployment cannot match.
Performance Benchmarks: What the Numbers Actually Show #
The Nature paper presents extensive benchmarking across multiple interaction types and test sets. The headline results demonstrate consistent improvement over specialized state-of-the-art methods—a remarkable achievement for a generalist model competing against task-specific tools.
The primary benchmark suite includes:
- PoseBusters (428 protein-ligand structures): Released to the PDB in 2021 or later, not in training data
- Recent PDB evaluation set (8,856 complexes): Released between May 2022 and January 2023
- CASP15 RNA targets (10 structures): Critical Assessment of Structure Prediction competition set
- Antibody-antigen complexes (65 interface clusters): Low-homology therapeutic-relevant systems
| Metric | AlphaFold 3 Result | Comparison Baseline | Statistical Significance |
|---|---|---|---|
| PoseBusters success (pocket RMSD < 2Å) | 76.6% | Vina: ~50%, Gold: ~45% | P = 2.27 × 10−13 vs. Vina |
| Protein-protein interface (DockQ > 0.23) | Improved rate | AlphaFold-Multimer v2.3 | P = 1.81 × 10−18 |
| Protein-nucleic acid interface | Higher accuracy | RoseTTAFold2NA | P = 7.28 × 10−12 |
| Antibody-antigen (high accuracy) | 42.4% @ 1,000 seeds | AlphaFold-Multimer: 25.4% | P = 0.009 |
| Protein monomer LDDT | Significantly improved | AlphaFold-Multimer v2.3 | P = 1.74 × 10−34 |
These numbers demonstrate that AlphaFold 3 is not merely comparable to specialized tools—it consistently outperforms them across interaction types. The statistical significance values (P < 0.001 for most comparisons) leave little doubt about the reliability of these improvements.
CASP15 and Recent Evaluations #
The CASP15 RNA targets provide a particularly informative benchmark because they represent blind prediction—structures determined after the prediction deadline. AlphaFold 3 achieves higher average performance than RoseTTAFold2NA and AIchemy_RNA (the top AI-based submission in CASP15) on the common subset of predictions.
However, the paper notes that AlphaFold 3 does not reach the performance of the best human-expert-aided CASP15 submission (AIchemy_RNA2). This gap between automated prediction and human-assisted prediction suggests room for improvement—either through architectural advances or through better integration of biological knowledge into the prediction process.
The more recent PDB evaluation set—8,856 structures released between May 2022 and January 2023—provides a larger-scale assessment of generalization. The structures span the full diversity of PDB deposits: monomers, homomers, heteromers, protein-nucleic acid complexes, ligand-bound structures, and post-translationally modified proteins.
Evaluation methodology matters here. The researchers applied strict homology filtering: individual chains with greater than 40% sequence identity to training set chains were excluded, as were interfaces where both polymers had homologous training examples. This ensures the reported accuracies reflect genuine generalization, not memorization of similar training structures.
The "cluster-weighted" scoring approach is also notable. Rather than counting each structure equally, the researchers cluster similar sequences and weight by inverse cluster size. This prevents over-representation of common protein families (like immunoglobulins or kinases) from inflating accuracy metrics. The resulting scores reflect performance across protein diversity, not just performance on the most common types.
Where AlphaFold 3 Still Struggles #
Despite its strengths, AlphaFold 3 has documented limitations that researchers must understand. The Nature paper is admirably explicit about these failure modes, providing guidance on when predictions should be treated with skepticism.
Stereochemical violations occur in a minority of predictions. The paper reports two main classes:
- Chirality violations (4.4% of predictions): The model occasionally produces molecules with incorrect stereochemistry at chiral centers, despite receiving correct reference structures as input features. The PoseBusters ranking includes a chirality penalty, but some violations persist.
- Atomic clashes: Some predictions contain overlapping atoms, particularly in large protein-nucleic acid complexes with more than 2,000 total residues and 100+ nucleotides. The ranking includes clash penalties but does not eliminate all cases.
Hallucinations in disordered regions are a known issue with diffusion-based models. Unlike AlphaFold 2, which produced distinctive ribbon-like structures in unstructured regions, AlphaFold 3 sometimes generates plausible-looking but spurious structure. The cross-distillation training from AlphaFold-Multimer predictions mitigates this but does not eliminate it. Fortunately, these hallucinated regions are typically marked by very low pLDDT scores, alerting researchers to the problem.
Conformational state limitations are perhaps the most biologically significant issue. AlphaFold 3 predicts static structures similar to those in the PDB. It does not model:
- Dynamic conformational ensembles
- Allosteric transitions between states
- Apo/holo conformational switching
The paper documents a striking example: cereblon, an E3 ubiquitin ligase, adopts an open conformation when unbound and a closed conformation when bound to ligands. AlphaFold 3 predicts the closed conformation for both states, apparently unable to capture the ligand-dependent conformational switch without explicit modeling of the binding event.
Accuracy dependence on MSA depth persists from AlphaFold 2. Proteins with shallow multiple sequence alignments—either because they are rarely sequenced or because they evolve rapidly—are predicted with lower accuracy. This creates a systematic bias: well-studied model organism proteins are predicted more accurately than exotic or poorly characterized sequences.
Technical Limitations and Honest Caveats #
AlphaFold 3 is a powerful tool, not a magic solution. Understanding its boundaries prevents misuse and sets appropriate expectations for what it can and cannot deliver to biological research.
The limitations fall into three categories: technical constraints of the model, scope restrictions of the training data, and fundamental limitations of structure prediction as an approach to biology.
Technical Constraints:
| Limitation | Impact | Mitigation |
|---|---|---|
| Static structure prediction | Cannot model dynamics, folding pathways, or conformational ensembles | Use experimental methods or specialized MD simulations for dynamics |
| Training data recency | No structures deposited after September 2021 in training | Predictions for recently evolved proteins may be less accurate |
| Chirality violations | 4.4% of ligand predictions have incorrect stereochemistry | Visual inspection and chirality checking in downstream analysis |
| Atomic clashes | Occasional in large complexes | Ranking with clash penalties reduces but does not eliminate |
| Hallucination in disordered regions | May generate spurious structure where none exists | Check pLDDT scores; low confidence indicates likely disorder |
Training Data Limitations:
AlphaFold 3 learns from the Protein Data Bank, which means it learns the biases and limitations of structural biology as a field. Membrane proteins remain under-represented relative to their biological importance. Intrinsically disordered proteins are rarely crystallized and thus poorly represented. Conformational heterogeneity is filtered out by the crystallization process.
The training cutoff (September 2021 for most evaluations, September 2019 for PoseBusters) means recently evolved proteins—like those from rapidly evolving viral strains—may be predicted less accurately. The model has not seen the structures that emerged from the structural biology pipeline in the past three years.
Fundamental Limitations:
Structure prediction is not biology prediction. Knowing the 3D coordinates of atoms in a protein complex does not tell you:
- Whether the complex forms in vivo
- The kinetics of formation and dissociation
- Post-translational modification states
- Cellular localization and trafficking
- Functional consequences of the interaction
AlphaFold 3 provides structural hypotheses that require experimental validation. It accelerates the hypothesis generation phase of research but does not replace the need for biological validation through assays, genetics, and physiology.
Dynamic Conformations and Predictive Uncertainty #
AlphaFold 3 predicts single static structures, not the conformational ensembles that many proteins actually populate. This is a fundamental limitation of the approach, not merely an implementation detail.
The diffusion process generates structures through iterative denoising from random initialization. Different random seeds produce different outputs, but these outputs do not represent a Boltzmann ensemble of thermally accessible conformations. They represent alternative predictions that the model finds plausible, with no guarantee that their distribution matches physical reality.
This matters for function. Many proteins operate as molecular machines—switching between conformations to transmit signals, transport molecules, or catalyze reactions. A static structure prediction, however accurate, captures only one snapshot of what may be a dynamic functional cycle.
The paper explicitly notes this limitation: "multiple random seeds for either the diffusion head or the overall network do not produce an approximation of the solution ensemble." Researchers who need ensemble information must look to other methods:
- Molecular dynamics simulations to explore conformational space
- Enhanced sampling techniques like metadynamics or replica exchange
- Experimental methods like NMR, DEER, or single-molecule FRET that capture dynamics
- MSA subsampling methods that have been developed for AlphaFold 2 and may transfer to AlphaFold 3
Predictive uncertainty is another nuanced topic. AlphaFold 3 provides confidence scores (pLDDT, ipTM) that correlate with accuracy, but these are epistemic uncertainty estimates—"how much does the model know?"—not aleatoric uncertainty about physical reality. A high-confidence prediction might be confidently wrong if the model has learned a systematic bias. A low-confidence prediction might reflect genuine structural disorder or simply limited training data for that protein family.
The Training Data Question #
AlphaFold 3's capabilities and limitations both stem from its training data: the Protein Data Bank. Understanding the PDB's biases helps interpret AlphaFold 3's predictions appropriately.
The PDB is not a random sample of biological structures. It reflects:
- Crystallographic tractability: Proteins that crystallize well are over-represented
- Solubility requirements: Membrane proteins and aggregates are under-represented
- Research interests: Human proteins, pathogen proteins, and model organism proteins dominate
- Technical evolution: Recent structures are higher resolution and better validated
The training pipeline described in the Nature paper uses structures deposited before September 30, 2021, with templates from a September 28, 2022 snapshot. The Chemical Components Dictionary—defining small molecules and ligands—was downloaded October 19, 2023.
This creates several specific biases:
Temporal cutoff: Protein families that have seen structural biology attention since 2021 are under-represented. Newly characterized viral proteins, for example, may not be well-captured.
Ligand diversity: The PDB represents drug-like chemical space unevenly. Well-funded therapeutic areas (oncology, metabolic disease) have richer ligand structure coverage than neglected diseases.
Protein size: Very large complexes (>5,000 residues) are rare in the PDB, limiting training signal for mega-complexes like nuclear pores or entire ribosomes.
Resolution variation: Training on both high-resolution crystal structures and lower-resolution cryo-EM maps means the model learns to reproduce the noise and artifacts of lower-quality experimental data alongside genuine structural signal.
The researchers address some of these concerns through careful evaluation set construction—filtering by homology, clustering by sequence similarity, and quality-filtering ligand structures. But the fundamental constraint remains: AlphaFold 3 predicts structures that look like PDB structures because it was trained to reproduce PDB structures.
This creates a "reality distortion field" where the model may systematically mispredict structures for systems that differ from typical PDB entries. Extremely large complexes, unusual chemistries, or novel folding topologies may be handled poorly—not because the architecture is incapable, but because the training data provides limited signal for these cases.
What This Means for AI in Life Sciences #
AlphaFold 3 marks an inflection point for AI in biology: the transition from single-task prediction to unified systems modeling. The broader pattern is clear—artificial intelligence is moving from automating individual analytical tasks to modeling the complex interactions that define biological function.
This has implications beyond structural biology:
For computational biology: The success of AlphaFold 3 validates the "end-to-end learning" approach. Rather than building modular pipelines (MSA generation → feature extraction → structure prediction → refinement), the model learns directly from sequence to structure. This suggests that other complex biological prediction problems—gene regulation, metabolic flux, cellular signaling—may yield to similar unified approaches.
For experimental biology: The relationship between computation and experiment is shifting. Computation becomes the first step, generating hypotheses that guide experimental design. Experiments become validation and discovery of the exceptional cases that models miss, rather than primary structure determination for routine cases.
For drug discovery: The integration of AI into the pharmaceutical industry accelerates. Isomorphic Labs' partnerships with Lilly and Novartis are early indicators of a broader trend: AI-native drug discovery companies partnering with traditional pharmaceutical companies to improve success rates. AlphaFold 3's ligand prediction capabilities make structure-based design viable for targets that previously lacked structural characterization.
For AI research: The architectural innovations in AlphaFold 3—particularly the Pairformer and diffusion module—suggest design patterns for other geometric deep learning problems. The success of simplified attention patterns and generative diffusion for 3D structure may transfer to materials science, chemistry, and robotics.
Democratization of Structural Biology #
AlphaFold 3 and the free server lower the barriers to entry for structural biology research. Previously, meaningful structural work required either experimental infrastructure (crystallography, cryo-EM) or computational resources (GPU clusters for running AlphaFold 2 locally). Now, any researcher with a web browser can generate structural hypotheses.
The implications for global science distribution are significant:
- Resource-limited institutions gain access to state-of-the-art structure prediction without capital investment
- Researchers in developing countries can participate in structural biology without expensive infrastructure
- Small labs can compete with well-funded structural genomics consortia on prediction tasks
- Interdisciplinary researchers from chemistry, physics, and engineering can incorporate structural insights without becoming structural biology specialists
This is not merely about cost savings. It is about cognitive accessibility. When structure prediction requires weeks of setup, troubleshooting, and computational management, only specialists can engage with it. When it requires pasting a sequence into a web form, the cognitive barrier drops and structural thinking becomes available to biologists of all stripes.
The free server model—non-commercial use only—strikes a balance between broad scientific access and commercial value capture. DeepMind and Isomorphic Labs are not charity organizations; they have made strategic choices about where to offer free access (basic research) and where to charge (drug discovery). This boundary is imperfect and will likely generate friction, but it represents a reasonable attempt to serve both scientific and commercial objectives.
The longer-term question is sustainability. Free services can be withdrawn, throttled, or monetized. Researchers building workflows around AlphaFold Server should consider what happens if access terms change or if the service experiences outages. The partial code release—allowing academic self-hosting without full model weights—provides some insurance but not complete independence.
The Next Research Frontier #
AlphaFold 3 establishes a foundation for the next phase of AI in structural biology. Several directions emerge naturally from the current capabilities:
Real-time and dynamic prediction: The static structure limitation is the most obvious frontier. Future models that predict conformational ensembles, folding pathways, and dynamic transitions would address a major biological need. Combining diffusion models with ideas from molecular dynamics or with learned force fields could enable this.
Integration with large language models: The parallel development of protein language models (ESM, ProGen) and structure prediction models suggests obvious synergies. A unified model that processes both sequence and structure through a common representation could achieve better few-shot prediction for novel folds and could generate sequences for desired structures (the inverse folding problem).
Cellular context: AlphaFold 3 predicts isolated molecular complexes, but biology occurs in crowded cellular environments. Predicting how structures change in the presence of membranes, crowding agents, and post-translational modification states would bring predictions closer to physiological reality.
Ligand generality: The current server limitation to curated ligands is a practical constraint, not a technical one. The Nature paper demonstrates prediction of arbitrary molecules via SMILES. Full deployment of this capability would enable structure-based drug design for any molecule a chemist can draw.
Accuracy for hard cases: The antibody-antigen result—continuing improvement with 1,000 seeds—suggests that sampling and ranking strategies remain important. Better confidence estimation, active learning from experimental data, and uncertainty-guided sampling could push accuracy on the hardest prediction targets.
The Nature paper's discussion section offers a measured view: "We expect that structural modelling will continue to improve not only due to advances in deep learning but also because continuing methodological advances in experimental structure determination...will provide a wealth of new training data." This symbiotic view—computational and experimental progress reinforcing each other—represents the most likely path forward.
AlphaFold 3 is not the end of the story. It is the foundation upon which the next decade of computational structural biology will be built.
Frequently Asked Questions #
What is AlphaFold 3 and how is it different from AlphaFold 2? #
AlphaFold 3 is a unified biomolecular structure prediction system that extends beyond proteins to model proteins, DNA, RNA, small molecules, ions, and post-translational modifications in a single framework. AlphaFold 2 predicted protein structures in isolation; AlphaFold 3 predicts how these molecules interact in complexes. The architecture replaces AlphaFold 2's Evoformer with a simpler Pairformer module and uses a diffusion-based structure generator instead of the invariant point attention structure module. This enables AlphaFold 3 to handle arbitrary chemical compositions—essential for modeling ligands, nucleic acids, and modified residues that AlphaFold 2 could not process.
When was AlphaFold 3 published in Nature? #
AlphaFold 3 was published in Nature on May 8, 2024. The publication coincided with the launch of the free AlphaFold Server at alphafoldserver.com, making the technology immediately accessible to researchers worldwide. The paper—titled "Accurate structure prediction of biomolecular interactions with AlphaFold 3"—documents the architecture, training methodology, and extensive benchmarking across protein-ligand, protein-nucleic acid, antibody-antigen, and other interaction types.
Is the AlphaFold 3 server free to use? #
Yes, the AlphaFold Server at alphafoldserver.com is free for non-commercial research use. The service requires only a Google account and provides access to AlphaFold 3's core prediction capabilities through a web interface. However, strict terms apply: outputs cannot be used commercially, cannot be used to train competing machine learning models, and cannot be fed into docking or screening tools. For commercial drug discovery applications, Isomorphic Labs—DeepMind's spinout company—provides licensed access to the full technology stack.
What types of biomolecules can AlphaFold 3 predict? #
AlphaFold 3 predicts the joint structure of complexes containing proteins, DNA, RNA, small molecule ligands, ions, and chemically modified residues. This unified scope is the primary advance over previous methods that specialized in single molecular types. The free server supports a curated subset of these capabilities—proteins, nucleic acids, limited ligands, and glycosylation—while the full model demonstrated in the Nature paper handles arbitrary small molecules specified by SMILES strings and a broader range of post-translational modifications.
How accurate is AlphaFold 3 for drug discovery applications? #
AlphaFold 3 achieves substantial accuracy improvements over existing methods for drug-relevant applications. On the PoseBusters benchmark of 428 protein-ligand structures, it achieves 76.6% success rate (pocket-aligned RMSD < 2Å), significantly outperforming classical docking tools like Vina and Gold. For antibody-antigen prediction, it substantially improves over AlphaFold-Multimer v2.3 (P = 6.54 × 10−5). These accuracies make it suitable for hit identification, target validation, and lead optimization in structure-based drug design workflows.
What are the main limitations of AlphaFold 3? #
AlphaFold 3 has documented limitations in stereochemistry, dynamic conformation prediction, and disordered regions. The model produces chirality violations in approximately 4.4% of ligand predictions and occasional atomic clashes in large complexes. It predicts static structures and cannot capture conformational ensembles or dynamic transitions between states. It may hallucinate structure in intrinsically disordered regions—though these are typically marked by low confidence scores. Accuracy also depends on multiple sequence alignment depth; proteins with few evolutionary relatives are predicted less reliably.
How does AlphaFold 3 handle protein-ligand interactions? #
AlphaFold 3 uses a diffusion-based generative model that learns the joint distribution of protein and ligand coordinates. Unlike traditional docking that requires predefined binding pockets and samples ligand conformations within them, AlphaFold 3 generates the bound complex directly from protein sequence and ligand SMILES specification. The diffusion process captures both protein flexibility (induced fit) and ligand conformational preferences without explicit pocket specification. This approach outperforms physics-based docking on benchmarks while operating without privileged pocket information.
What is the relationship between DeepMind and Isomorphic Labs? #
Isomorphic Labs is a DeepMind spinout company founded to commercialize AlphaFold technology for drug discovery. Demis Hassabis serves as CEO of both entities. DeepMind pursues open science—publishing research, maintaining free resources like the AlphaFold Server, and releasing code for academic use. Isomorphic Labs applies the technology commercially through partnerships with pharmaceutical companies. In January 2024, Isomorphic announced deals with Eli Lilly ($45 million upfront, up to $1.7 billion total) and Novartis ($37.5 million upfront, up to $1.2 billion total) for AI-driven drug discovery collaborations.
Can AlphaFold 3 replace experimental methods like X-ray crystallography? #
AlphaFold 3 complements but does not fully replace experimental structure determination. For routine cases—soluble proteins with good homology to known structures—it often produces accurate predictions that reduce the need for experiments. However, crystallography and cryo-EM remain essential for novel folds, validation of predictions, membrane proteins, and capturing dynamic conformational states that AlphaFold 3 cannot predict. The relationship is shifting: computation becomes the first step for hypothesis generation, while experiments focus on validation and frontier cases beyond current predictive capabilities.
How do I access the AlphaFold 3 server today? #
Navigate to alphafoldserver.com and sign in with a Google account. The interface accepts protein sequences (FASTA format), DNA/RNA sequences, and selections from a curated ligand library. Typical predictions complete in 2-10 minutes depending on complex size. Results include 3D structure visualization, confidence scores (pLDDT, pTM, ipTM), predicted aligned error matrices, and downloadable structure files in mmCIF format. For programmatic access or custom ligand support, DeepMind has indicated that inference code will be released for academic use, though without full model weights.
Closing: The Broader Pattern AI Builders Should Recognize #
AlphaFold 3 exemplifies a pattern I see repeatedly in production AI systems: the unification of specialized pipelines into end-to-end learned models. Where previous approaches chained together modular tools (MSA generation, feature extraction, structure prediction, refinement), AlphaFold 3 learns directly from sequence to structure. The result is not just better accuracy—it is a radically simpler system that generalizes across molecular types that previously required separate handling.
This pattern applies beyond structural biology:
- Workflow automation moves from scripted pipelines to AI agents that learn task structure
- Content generation moves from template-based systems to generative models that learn the distribution of effective content
- Decision systems move from rule engines to learned predictors that capture tacit knowledge from historical outcomes
The engineering lesson is clear: when you have sufficient data and compute, end-to-end learning often beats carefully engineered pipelines. The architecture should follow the data, not the other way around.
For organizations building with AI, AlphaFold 3 also demonstrates the value of managed platforms over self-hosted complexity. The AlphaFold Server makes state-of-the-art capability accessible to researchers who lack GPU infrastructure or ML engineering expertise. The pattern—research systems → open databases → managed services—creates value while managing operational complexity.
If you're building AI-powered systems for your organization—whether automating workflows, structuring knowledge pipelines, or integrating models into production—I can help architect and implement the infrastructure. Book an AI automation strategy call to discuss how to apply these patterns to your specific domain.
The biomolecular revolution is here. The question for builders is: what analogous revolution can you enable in your field?
Related Reading #
- Claude 3.5 Sonnet: Anthropic's Fastest Model Yet — How Anthropic's parallel model development strategy mirrors DeepMind's AlphaFold release pattern
- Google I/O 2024: The AI Count and Project Astra — Google's broader AI infrastructure push that includes the AlphaFold Server
- The GPT-4o Launch: Omni-Model Goes Free Tier — The parallel trend of making frontier AI capabilities freely accessible
- Building AI Agent Systems That Actually Work — Architectural patterns for production AI systems that apply lessons from AlphaFold's unified model approach
Related Posts

Google I/O 2026 Action List: How I Prompted Gemini 3.5 Flash and Antigravity Workflows
Google I/O 2026 just reset the AI tooling landscape. Here's the 9-action checklist for builders who want to ship this week, not just watch the keynote.

Anthropic vs. OpenAI vs. Google: The State of the Frontier in May 2026
A head-to-head breakdown of the three AI giants in May 2026: Claude Opus 4.6, GPT-5.3 and 5.4, Gemini 3.1 Pro. Real specs, real pricing, and what actually matters for builders.

Kimi K2 Open Weights: How I Prompted Moonshot's Frontier Model for Agentic Tool Use
How I direct Kimi K2 by Moonshot AI for agentic workflows, long-context tool calling, and workflow automation. A 1 trillion parameter MoE model with competitive benchmarks at 5-17x lower cost than GPT-5 and Claude.